In 2001 publiceerden Marcus Raichle en collega’s, verbonden aan Washington University School of Medicine, een serie Positron Emission Tomography (pet)-experimenten (Raichle et al., 2001; Shul-man et al., 1997). Deze techniek stelt ons in staat te kijken naar het metabolisme in ons brein.
Interessant aan deze experimenten was dat wanneer mensen in rust waren het metabolisme door het hele brein volledig homogeen was, zoals gemeten met de oxygen extraction fraction: de ratio tussen energietransport en energieverbruik. Op basis hiervan stelden deze onderzoekers vast dat passieve rust de normale – meest energie-efficiënte – bewuste toestand van het brein is. Deze vormt met andere woorden onze default mode van het brein: het brein is dan metabolisch gezien volledig in evenwicht. Opvallend is dat wanneer we weer een taak gaan uitvoeren dit metabolische evenwicht verdwijnt.
Naast deze metabolische bevindingen kan met een andere neuro-imaging techniek, fMRI – die is gebaseerd op bloedtoevoer naar actieve hersengebieden – in kaart worden gebracht welke hersengebieden bij onze bewuste rusttoestand daadwerkelijk actief zijn.
Een netwerk
Een groot aantal fmri-studies hebben inmiddels een rustnetwerk in kaart gebracht bestaande uit de posterior midline region (posterior cingulate en mediale parietale gebieden), laterale pariëtale gebieden en de mediale prefrontale cortex (Buckner et al., 2008 en figuur 1; Davanger, 2015). Dit netwerk staat inmiddels bekend als het default mode netwerk (DMN) of korter ‘default netwerk’ dan wel ‘standby netwerk’. Dit netwerk is actiever tijdens rust dan tijdens het uitvoeren van veeleisende extern-gerichte cognitieve taken.
Het DMN is een netwerk van verschillende onderling verbonden hersengebieden bestaande uit de frontale en pariëtale kwabben die ver uiteenliggen. Het gaat dus niet om één speciale plek in de hersenen. De verschillende gebieden staan met elkaar in contact via het zenuwnetwerk en de communicatie daarbinnen gebeurt via de zogeheten ‘witte stof’ (myeline), die om de uitlopers (neurieten) van zenuwcellen heen zit. De myeline vormt een soort schacht, die zorgt voor een betere gelei-ding van de zenuwimpulsen.
Het DMN wordt verondersteld een gebied te zijn waar heel veel neurieten samenkomen. Een zenuwcentrum dus waarin allerlei soms vér uiteenliggende hersengebieden contact met elkaar maken. Een Amerikaans samenwerkingsverband van onderzoekers vond dat de functionele verbindingen tussen de hersendelen binnen het DMN op de leeftijd van zeven tot negen jaar vergeleken bij volwassenen nog zwak zijn, doordat de myelinisering nog gaande is (Fair et al. 2007). Waarschijnlijk zijn kinderen daardoor minder tot introspectie in staat. Ook het ontstaan van zelfbewustzijn bij kinderen – ‘ik ben ik’ – zal vermoedelijk verband houden met de ontwikkeling van het DMN .
Gericht op de buitenwereld
In Amerikaanse publicaties over dit netwerk wordt vaak mind-wandering gebruikt als conditie waaronder het DMN activiteit gaat vertonen. Christoff et al. (2016) stellen dat mind-wandering het best gezien kan worden als lid van een familie van geestestoestanden waartoe ook het creatieve denken en het dagdromen behoren. Een geestestoestand die wij allemaal kennen uit situaties waarin je niet druk bezig bent met het uitvoeren van opdrachten, het zoeken van dingen of het oplossen van problemen. Het zijn de momenten zoals wanneer je in de trein of auto wordt gereden en je het landschap aan je voorbij ziet trekken, of als je lang gedachteloos onder de douche staat of op een strandstoel ligt niets te doen.
De met behulp van fmri-scans waargenomen activiteit in het DMN gaat gewoonlijk gepaard met wat men is gaan samenvatten als random episodic silent thinking about one’s life – als men gedachten de vrije loop laat. Het DMN vertoont pas activiteit als de proefpersonen geen enkele taak te verrichten hebben, of wel een opdracht krijgen, maar één die totaal geen cognitieve inspanning vergt, zoals het zo ontspannen mogelijk kijken naar een gekleurd punt in het gezichtsveld. Bij het uitvoeren van opdrachten van de proefleider, zoals het maken van sommen of het oplossen van (verbale) raadsels, is men gericht op de buitenwereld en niet ‘naar binnen gericht op het eigen innerlijk’ zoals Swaab (2016) dat heel psychologisch in zijn laatste boek verwoordt. Het DMN is dan juist niet actief.
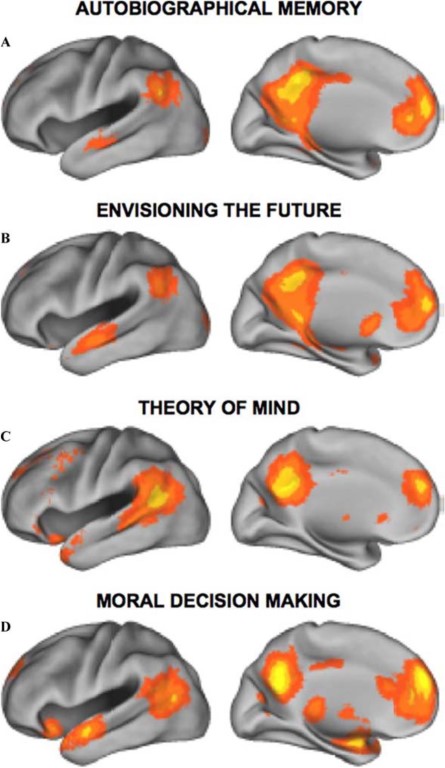
Interessant is dat volgens de ontdekkers en diverse latere onderzoekers het DMN niet alleen actief is tijdens rust, maar ook tijdens allerlei introspectieve bezigheden, zoals dagdromen, denken aan het verleden, denken aan de toekomst, mentale inbeelding en het maken van moeilijke morele beslissingen (Buckner et al. 2008 en figuur 2). Maar de geestestoestand waarin men bij proefpersonen de meeste activiteit in het DMN te zien krijgt, is als zij opdrachten krijgen die een beroep doen op inlevingsvermogen, op sociale cognitie dus (Mars et al. 2012; Salet et al., 2011). Daarom is men de bij het DMN betrokken gebieden the social brain gaan noemen. (Overigens publiceerde de neuropsycholoog Michael Gazzaniga in 1985 al een boek met als titel The social brain en als ondertitel Discovering the networks of the mind – dus al zo’n vijftien jaar vóór de ontdekking van het DMN).
Al snel na de ontdekking van het DMN is men ook bij (andere) apen gaan zoeken naar activiteit in dat social brain (Dunbar, 1998; 2012). Bij makaken die onder lichte verdoving werden gebracht, vonden onderzoekers interessante resultaten (Vincent et al. 2006). Met fmri stelden ze vast dat de hersengebieden die betrokken zijn bij het DMN groter zijn bij makaken die in grotere groepen leven – dus met meer sociale verbanden – en ook naarmate zij een hogere positie hebben binnen de leefgroep (Sallet et al., 2011).
Meditatieoefeningen
Een internationaal team van onderzoekers heeft het DMN onderzocht bij een ‘niet-directieve’ vorm van mediteren (ACEM), waarbij het laten afdwalen van de gedachten niet wordt tegengegaan. Dit in tegenstelling tot meditatievormen waarbij men zich moet concentreren. Van de ACEM-vorm van meditatie wordt verondersteld dat deze de psychische verwerking van (herinneringen aan) emotionele gebeurtenissen helpt bevorderen (Taylor et al., 2012). De proefpersonen waren ervaren beoefenaars van deze meditatievorm. Er waren twee experimentele condities. In de eerste conditie werd met fmri-metingen de activiteit in verschillende delen van het brein gemeten tijdens de standaard DMN in rustsituatie (resting state), en vergeleken met een situatie waarin op ontspannen wijze gemediteerd werd. In de tweede conditie werd de proefpersonen gevraagd zo geconcentreerd mogelijk te mediteren – bijvoorbeeld door intensief op de ademhaling te letten – en werden de resultaten daarvan vergeleken met die van de ontspannen meditatie uit de eerste conditie.
Wat bleek? In de eerste conditie werd tijdens het ACEM-ontspannen mediteren significant méér activiteit vastgesteld in hersengebieden die geassocieerd worden met mind-wandering, het ophalen van belevenissen uit het geheugen en de verwerking van emotionerende gebeurtenissen dan tijdens een standaard resting state-situatie. Vergeleken met de tweede conditie liet bij de ontspannen meditatie uit de eerste conditie vooral het gebied dat in verband gebracht wordt met het ophalen van gebeurtenissen uit het geheugen en de emotionele verwerking daarvan meer activiteit zien.
De onderzoekers concludeerden dat niet-restrictieve meditatietechnieken inderdaad helpen als het doel van het mediteren is emotionerende gebeurtenissen uit het verleden beter te kunnen ver-werken en de met die herinneringen verbonden stress te kunnen verlagen.
Theory of mind
Onderdelen van het DMN zijn niet alleen actief tijdens het denken aan onszelf, maar ook tijdens het ons verplaatsen in het denken van iemand anders, oftewel Theory of Mind (ToM). Ook dit is een introspectief proces. In een vroege studie hebben Saxe & Kanwisher (2003) gekeken naar de neurale correlaten daarvan. Zij vonden dat binnen de laterale pariëtale cortex – de temporo-parietelal junction (tpj) – dat de scheiding vormt tussen temporaal en pariëtaal, in het bijzonder betrokken was tijdens het denken over de inhoud van het denken van een ander. De tpj reageerde niet op niet-sociale verhalen, maar wel sterk op sociale verhalen. Verder werd het ook meer actief wanneer mensen verhalen lazen over de geestestoestand van een personage ten opzichte van verhalen die de fysieke details van mensen beschreven. Zo lijkt de tpj belangrijk te zijn voor het begrijpen van andere mensen.
Saxe & Kanwisher (2003) hebben ook gekeken bij kinderen naar de ontwikkeling van ToM. Activiteit van DMN-hersengebieden werd gemeten terwijl kinderen (tussen vijf en elf jaar oud) en volwassenen luisterden naar de beschrijvingen van geestestoestanden van personages, in vergelijking met beschrijvingen van hun fysieke eigenschappen. Bij de kinderen werden de reacties vanuit de tpj steeds sterker tijdens het luisteren naar de verhalen over geestestoestanden, maar niet wanneer de verhalen gingen over het uiterlijk van mensen. Functionele activiteit in de rechter tpj was gerelateerd aan de prestaties van de kinderen op een moeilijke ToM-taak. Dus dit onderdeel van het DMN lijkt met name belangrijk te zijn bij de ontwikkeling van kinderen in het zich kunnen verplaatsen in andermans situatie.
Verschillen binnen het DMN
Het DMN vormt een sterk gekoppeld netwerk, maar is niet helemaal homogeen. Er zijn sterkere of minder sterkere verbindingen met verschillende onderdelen van het DMN en andere netwerken of hersengebieden. De laterale en mediale pariëtale componenten zijn meer betrokken bij algemene interne processen, zoals geheugen. Met name de posterior cingulate heeft een sterker verband met de mediaal temporaal kwab die belangrijk is voor het geheugen (zie figuur 3). De mediale prefrontale component aan de andere kant wordt vooral geassocieerd met self-related processing en heeft een sterkere koppeling met orbitofrontale gebieden, belangrijk voor emotionele verwerking.
De klinische praktijk
Ook vanuit klinisch oogpunt is er interesse in het DMN. Zo is men in de biologische psychiatrie gaan onderzoeken of het op het innerlijk gericht zijn van het DMN ook excessief kan worden en dan een verklaring zou kunnen zijn voor het langdurig blijven piekeren (rumineren) zoals bij Major Depressive Disorders (Dutta et al. 2014; Hamilton et al, 2015). Maar stevige conclusies zijn er nog niet. Bij andere klinische stoornissen is wel duidelijk verband aangetoond met het aangedaan zijn van het DMN. Hier bespreken we kort een paar bevindingen van de rol van het DMN voor Alzheimer, schizo-frenie en psychopathie.
De ziekte van alzheimer
Een van de belangrijkste symptomen bij Alzheimer dementie-patiënten zijn problemen met het geheugen (Buckner et al. 2008). Zoals vermeld speelt het DMN een belangrijke rol in het geheugen als intern proces. Aan de andere kant vereist het opslaan van informatie) als extern proces dat het DMN uitgeschakeld wordt (de-activatie; Daselaar, et al. 2004; Daselaar et al., 2009; Huijbers et al 2009; Huijbers et al. 2012). Er is gevonden dat Alzheimer dementie-patiënten de deactivaties tijdens het opslaan van informatie al in een heel vroeg stadium minder laten zien ten opzichte van jongeren en gezonde ouderen. Deze bevinding kan een vroegere marker opleveren van de ziekte dan de veranderingen die optreden in de hippocampus, het gebied dat het meest met Alzheimer Dementie geassocieerd wordt. Bijvoorbeeld bij jongeren met het APOE4 gen – een belangrijke voorspeller voor het later ontwikkelen van Alzheimer-dementie – is gevonden dat er verminderde de-activaties optreden tijdens het opslaan ten opzichte van jongeren zonder het gen, maar dat er geen verschillen zijn in de hippocampus (Persson et al., 2008).
Verder is dit gebrek aan deactivatie tijdens het opslaan, ten opzichte van activatie tijdens herinneren, gekoppeld aan de accumulatie van amyloide dat ook gezien wordt als belangrijke voorspeller voor het ontwikkelen van Alzheimer-dementie. Ook hier is er geen relatie met hippocampale afwijkingen gevonden (Huijbers, Vannini et al. 2012; Vannini, Hedden et al. 2013). Deze bevindingen gaan samen met pet-studies die een afname laten zien in PMR metabolisme – zonder hippocampale afwijkingen – bij patiënten met de diagnose Mild Cognitive Impairment. Dit laatste is een conditie voor een verhoogde kans op het ontwikkelen van Alzheimer-dementie (Nestor et al., 2014).
Het lijkt er dus op dat DMN deactivaties tijdens het opslaan van informatie een betere marker opleveren van vroege stadia van Alzheimer-dementie dan hippocampale afwijkingen. Deze vroege veranderingen in het DMN kunnen een belangrijke factor worden voor het ontwikkelen van effectieve therapieën voor het voorkomen van deze vorm van dementie.
Schizofrenie
Er zijn ook duidelijke afwijkingen gevonden bij schizofreniepatiënten. Om te kijken naar hoe goed de DMN-componenten met elkaar verbonden zijn tijdens rust kan gebruik gemaakt worden van functionele connectiviteitsmethoden, zoals seed-based connectivity (Vincent et al., 2006) of independent component analysis (Egolf et al. 2004). Tijdens rust laten schizofreniepatiënten duidelijk een verminderde koppeling zien tussen de gebieden die het DMN vormen dan gezonde controles (Whitfield-Gabrieli et al., 2009). Hieruit zou kunnen worden afgeleid dat hun interne ‘huishouding’ niet goed georganiseerd is en de mate van DMN-koppeling kan bijvoorbeeld weer gebruikt worden als maat met betrekking tot hoe goed een bepaalde therapie of medicijn aanslaat. Dit kan weer nuttige consequenties hebben voor de verdere behandeling van een patiënt.
De studie van Zhou et al. (2016) is een goed voorbeeld van individuele verschillen, binnen de populatie gediagnosticeerd als schizofreen, met betrekking tot het DMN. De cognitieve functie bij schizofrenie patiënten varieert van betrekkelijk intact tot ernstig aangetast. Deze studie keek naar het deactivatie patroon van het DMN tijdens het uitvoeren van een werkgeheugentaak tussen schizofreniepatiënten met en zonder verminderde cognitieve functie. De groep met cognitieve gebreken vertoonde verminderde de-activatie in de mediale prefrontale cortex en mediale pariëtale cortex.
Dus: verstoorde DMN-activiteit lijkt op te treden bij schizofreniepatiënten met een verminderde cognitieve functie. Dit zijn slechts twee voorbeelden, die duidelijk aangeven dat het belangrijk is de rol van DMN-disfunctie in ogenschouw te nemen bij het diagnosticeren en behandelen van schizofrenie patiënten op individuele basis.
Psychopathie
Tot slot een onderzoek waar ook de eerder genoemde Gazzaniga bij betrokken was (Freeman et al. 2015). Gevangenen werden door psychiaters en psychologen beoordeeld op eigenschappen als empathie, spijt, antisociaal gedrag, overmatige zelfgerichtheid en gebrek aan bekommernis om anderen. Twee groepen werden gevormd van elk 22 personen, de ene groep van hoog scorende mensen op de psychopathie-index, en een andere groep van laag scorende. De resultaten van fmri-scans in een resting state na een taakgerichte periode waren dat de activiteit in het DMN lager was in de hoog psychopathische groep – met name in gebieden die in het algemeen in verband gebracht zijn met affectie-psychopathische eigenschappen, waaronder een gebrek aan mededogen met anderen.
Besluit
Het DMN leek aanvankelijk in een passieve toestand te verkeren. Inmiddels is men ervan overtuigd geraakt dat dit alleen schijnbaar zo is. Naast de activiteit van het brein in het omgaan met de buitenwereld is er ook een voortdurend actieve interne wereld. Bijvoorbeeld bestaande uit Rapid Episodic Silent Thinking (Andreasen et al. 1995). Rust is – in tegenstelling tot slaap – geen passieve toestand, maar juist een heel actieve, waarbij veel energie wordt gebruikt. Die is ruim voorhanden, doordat het DMN is voorzien van de grootste bloedvaten in het brein. De default mode is de tonische activiteit in het DMN (Gusnard et al. 2001).
Verschillende groepen patiënten vertonen afwijkingen in het DMN. De diagnostiek daarvan kan meer inzicht geven in het verloop van ziekteprocessen en behulpzaam worden in de behandeling van die processen. Nadere bestudering van het DMN is dan ook zowel vanuit klinisch als cognitief oogpunt belangrijk.
Er is behoefte aan meer neuro-ontwikkelingspsychologisch onderzoek, speciaal bij adolescenten. En dan apart voor jongens en meisjes, een helaas nog ontbrekend onderscheid in het meeste DMN-onderzoek. Eveneens meer onderzoek is gewenst naar de effectiviteit van verschillende vormen van meditatie- en mindfulnesstrainingen: hoe meer DMN-activiteit tijdens rust, des te effectiever zij blijken te zijn.
Literatuur
- Buckner, R.L., Andrews-Hanna, J.R. & Schacter, D.L. (2008). The Brain’s Default Network: Anatomy, function, and relevance to Disease. Annals of The New York Academy of Sciences, 1124, 1-38.
- Buckner, R. L., Snyder, A. Z., Shannon, B. J., LaRossa, G., Sachs, R., et al. (2005). Molecular, structural, and functional characteriza-tion of Alzheimer’s disease: evidence for a relationship between default activity, amyloid, and memory. Journal of Neuroscience, 25, 7709–7717.
- Buytendijk, F.J.J.(1947). Het kennen van de innerlijkheid. Oratie als hoogleraar Algemene Psychologie aan de Universiteit Utrecht.Nijmegen,Dekker &van de Vegt (citaat p.1).
- Christoff, K., Irving, Z.C., Fox, K.C.R., Spreng, R.N. & Andrews-Hanna, J.R.(2016). Mind-wandering as spontaneous thought: a dynamic framework. Nature Neuroscience Review 6(17), 718-73.
- Daselaar, S.M., Prince, S.E. & Cabaza, R. (2004). When less means more: deactivations during encoding that predict subsequent memory. Neuroimage, 23, 3921-3927.
- Daselaar, S.M., Prince, S.E. & Dennis, N.A. (2009). Posterior midline and ventral pariental activity is associated with retrieval success and encoding failure. Frontiers in Human Neuroscience, 3. doi: 10.3389/neuro.09.013.2009
- Daselaar, S.M., Huijbers, W., Eklund, K., Moscovitch, M. & Cabeza,R. (2013). Resting-state functional connectivity of ventral parietal regions associated with attention reorienting and episodic recollection. Frontiers in Human Neuroscience, 7, 38.
- Davanger, S. (2015). The brain’s default network – what does it mean to us? The Meditation Blog, march 19, 1-3.
- Dunbar, R.I.M. (1998). The social brain hypothesis. Evolutionary Anthropology, 6, 178-190.
- Dunbar, R.I.M. (2012). The social brain meets neuroimaging. Trends in Cognitive Science, 16, 101-102.
- Dutta, A., McKie, S. & Deakin, J.F.W. (2014). Resting state networks in major depressive disorder. Psychiatry Research, 224(3), 139-151.
- Egolf, E.A., Kielh, K.A. & Calhoun, V.D. (2004). Group ICA of fMRI Toolbox (GIFT). Poster presented at the annual meeting of the Society for Biological Psychiatry, New York, NY.
- Fair, D.A., Cohen, A.L., Dosenbach, N.U.F., Church, J.A., Miezin, F.M. et al. (2007). The maturing architecture of the brain’s default network. Proceedings of the National Academy of Sciences of the United States of America, 105(10).
- Freeman, S.M., Scott, M., Clewett, D.V., Craig, M., Gazzaniga, M.S. et al. (2015) The posteromedial region of the default mode network shows attenuated task-induced deactivation of the DMN in psychopathic prisoners. Neuropsychology, 29(3), 493-500.
- Gazzanniga, M. (1985). The Social Brain, Discovering the Networks of the Mind. New York Basic Books.
- Gusnard, D.A., Akbudak, E., Shulman, G.L. & Rachel, M.E. (2001) Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proceedings of the National Academy of Sciences of the United States of America, 98, 4259-4264.
- Hamilton, J.P., Farmer, M., Fogelman, P. & Gotlib, I.H. (2015) Depressive Rumination, the Default-Mode Network, and the Dark Matter of Clinical Neuroscience. Biological Psychiatry, 78(4), 224-230.
- Huijbers, W., Pennartz, C.M., Cabeza, R. & Daselaar, S.M. (2009). When learning and remembering compete: a functional MRI study. Public Library of Science, Biology, 7, 1. e.1000011.
- Huijbers, W., Vannini, P., Sperling, R.A., Pennartz, C.M.A., Cabeza, R. & Daselaar, S.M. (2012) Explaining the encoding/retrieval flip: Memory-related deactivations and activations in the posteromedial cortex. Neuropsychologia 50(14), 3764-3774.
- Kohnstamm, D. (2002). Ik ben ik, de ontdekking van het zelf. Amsterdam: De Bezige Bij.
- Kohnstamm, D. (2016). De eigenander; een ik in mij. Arnhem: Uitgeverij Nieuwe Druk.
- Mars, R.B, Neubert, F.X., Noonan, M.A.P., Sallet, J., Toni, I. et al. (2012). On the relationship between ‘the default mode network’ and the ‘social brain’. Frontiers in Human Neuroscience, 6, 189. doi: 10.3389/fnhum.2012.00189
- Nestor, P.J., Scheltens, P.J. & Hodges, J.R. (2014) Advances in the early detection of Alzheimer’s disease. Nat Med 10 Suppl: S34-41.
- Persson, J., Lind, J., Larsson, A. & Nyberg, L. (2008) Altered deactivation in individuals with genetic risk for Alzheimer’s disease. Neuropsychologia 46(6), 1679-1687.
- Raichle, M.E., MacLeod, A.M., Snyder, A.Z., Powers, W.J., Gusnard, D.A. et al. (2001). A default mode of brain function. Proceedings of the Academy of Sciences of the United States of America, 98(2), 676-682.
- Salet, J., Mars, R.B., Noonan, M.P., Andersson, J.L. Jbabdi, S. et al. (2011). Social network size affects neural circuits in macaques. Science, 334, 697-700.
- Saxe, R. & Kanwisher, N. (2003). People thinking about thinking people: the role of the temporo-parietal junction in “theory of mind”. Neuroimage, 19(4), 1835-1842.
- Shulman, G.L., Fiez, J.A., Corbetta, M., Buckner, R.L., Miezin, F.M. et al (1997). Common blood flow changes across visual tasks II. Decreases in cerebral cortex. Journal of Cognitive Neuroscience 9(5), 648-663.
- Swaab, D. Ons creatieve brein (2016). Amsterdam: Atlas Contact Uitgeverij.
- Taylor, V.A., Dancault, V., Grant, J., Scavone, G. Breton, E. et al. (2012). Impact of meditation training on the default mode network during restful state. Social Cognitive and Affective Neuroscience, march 24., 4028-4030.
Thermenos, H.W. zie Whitfield-Gabriel et al. - Vannini, P., Hedden, T., Huijbers, W., Ward, A., Johnson, K.A. et al. (2013). The ups and downs of the posteromedial cortex: age- and amyloid-related functional alterations of the encoding/retrieval flip in cognitively normal adults. Celeb Cortex, 23(6), 1317-1328.
- Vincent, J.L., Snyder, A.Z., Fox, M.D., Shannon, J., Andrews, J.R. et al. (2006) Coherent spontaneous activity identifies a hippocampal-parietal memory network. Journal of Neurophysiology 96(6), 3517-3531.
Whitfield-Gabrieli, S., Thermenos, H.W., Milanovic, S., Tsuang, M.T., - Faraone, S.V. et al. (2009) Hyperactivity and Hyperconnectivity of the default network in schizophrenia and in the first-degree relatives of persons with schizophrenia. Proceedings of the National Academy of Sciences of the United States of America, 106(4), 1279-1284.
- Zhou, I, Weidan Pu, Jingjuan Wang, Haihong Liu, Guowei Wu et al. (2016) Inefficient DMN Suppression in Schizophrenia Patients with Impaired Cognitive Function but not Patients with Preserved Cognitive Function. Scientific Reports, 6, Article number 21657.


